Genus Camellia: A review of extant taxonomic systems using the latest morphological and molecular data
Orel G.*, Curry A.S.
Royal Botanic Gardens, Mrs Macquaries Road, Sydney, NSW 2000, Australia
e-mail: george.orel@rbgsyd.nsw.gov.au
Introduction
Comparisons between the taxonomic systems of Sealy (1958), Chang & Bartholomew (1984), Ming & Bartholomew (2007) and of the synopses of Ho (1991) and Gao et al (2005) indicate the existence of diametrical differences in approaches to the taxonomy of genus Camellia, thus producing differing taxonomic systems. Sealy’s taxonomic system divided Camellia into some 12 sections, which comprised 106 species, 24 of which were considered to be doubtful (Sealy 1958). Chang & Bartholomew after revising their taxonomic system are best known for their 1984 publication (Chang & Bartholomew 1984). In this work they divided genus Camellia into 4 subgenera and 22 sections with some 280 Camellia species. Ming & Bartholomew (2007) reduced the number of Camellia species to 119 and divided them into 2 subgenera and 14 sections.
The above-mentioned authors derived their respective data from observation of the species morphological characters. It should be noted that, in some instances, the authors worked with herbarium specimens only, some of which were not complete, i.e. the specimens lacked flowers and/or seed capsules. Indications are that some of the herbarium specimens used may have been misidentified and were not always referred to the type specimens (Sealy 1958, Ho 1991, Gao et al 2005, Ming & Bartholomew 2007).
Standard works that describe the morphology of the members of genus Camellia often exhibit a relative paucity of morphological data. As a rule the descriptors are relatively short and record only some of the morphological characters, whilst omitting characters that at the time of the description were not available or not considered. Only the basic morphology is described and many of the secondary details are not incorporated.
The fact that the results of a number of molecular studies often differ from the studies based on morphological data further complicates the endeavour to create a unified taxonomic system for genus Camellia. There can be no doubt that the molecular studies are of great help in resolving some of the problems in Camellia phylogeny. However, a system based entirely on molecular data would give us a very different perspective re the interspecific relationships in the genus. Thus a hybrid system may have to be considered.
In this work the analyses of some 47 Camellia species (52 accessions) were performed to highlight the incompatibility of the existing taxonomies.
Materials and Methods
Morphological and molecular characters of some 47 Camellia species (52 accessions) were analysed to indicate the interspecific status of known Camellia species and also of the nine newly discovered Camellia taxa from Viet Nam. All materials were collected by the authors of this work and the exact provenance details for each were withheld for conservation reasons.
Most of the 30 multiple state characters were gained from in situ observations. The assessments of herbarium materials were performed under laboratory conditions. A morphological matrix, where the absence of a character was denoted 0 and the presence 1, 2, 3, 4, 5 and 6, was constructed. Phylip 3.69, a computational program for inferring evolutionary phylogenies was used to generate the taxonomic trees. The general provenance, the morphological and population data for all Camellia species used are presented in Tables 1. and 2.
Genomic DNA was extracted from fresh or freeze-dried leaves, using a method similar to that of Dellaporta et al. (1983, as described by Wilkie, 1997), with the additional modification of including 1% PVP (polyvinyl pyrrolidone) in the extraction buffer (following Singh et al 1999, who report methods for DNA fingerprinting of commercially prepared tea). Further purification was effected by a diatomaceous earth binding procedure, adapted from the technique described by Gilmore et al. (1993).
Table 1. General provenance and population data for 47 Camellia species (52 accessions) from Viet Nam and China that were used in this paper
|
Taxon* |
Provenance |
Population notes |
|
C. dongnaiensis* |
Viet Nam |
Less than 20 individual plants collected |
|
C. luteocerata |
Viet Nam |
Small scattered population |
|
C. inusitata |
Viet Nam |
Small relatively protected population |
|
C. oconoriana |
Viet Nam |
Relatively large population |
|
C. sp. 698** |
Viet Nam |
Small scattered population |
|
C. sp. OCa** |
Viet Nam |
Population size unknown |
|
C. sp. OCb** |
Viet Nam |
Population size unknown |
|
C. sp. OCc** |
Viet Nam |
Population size unknown |
|
C. sp. CT5** |
Viet Nam |
Population size unknown |
|
C. impressinervis |
Viet Nam |
Population size unknown |
|
C. rubriflora |
Viet Nam |
Population size unknown |
|
C. vidalii |
Viet Nam |
Population size small |
|
C. gilbertii |
Viet Nam |
Recently discovered new population |
|
C. petelotii |
Viet Nam |
Population large (Tam Dao N. P.) |
|
C. petelotii |
Viet Nam |
Cultivated specimen |
|
C. petelotii |
P.R. China |
Cultivated specimen |
|
C. limonia |
Viet Nam |
Population size small |
|
C. flava |
Viet Nam |
Population relatively large |
|
C. rosmannii |
Viet Nam |
Small population, some in cultivation |
|
C. oleifera |
Viet Nam |
Cultivated specimen |
|
C. aurea |
P.R. China |
Population size unknown |
|
C. nitidissima |
P.R. China |
Cultivated specimen |
|
C. nitidissima var. microcarpa |
P.R. China |
Taxonomic status uncertain |
|
C. nitidissima var. nitidissima |
P.R. China |
Population size not known |
|
C. luteoflora |
P.R. China |
Population size not known |
|
C. tunhingensis |
P.R. China |
Population size not known |
|
C. pingguoensis |
P.R. China |
Population size not known |
|
C. pingguoensis |
P.R. China |
Cultivated specimen |
|
C. salicifolia |
P.R. China |
Cultivated specimen |
|
C. pitardii var. alba |
P.R. China |
Cultivated specimen |
|
C. sp. (Halong Bay) |
Viet Nam |
Population size unknown |
|
C. sp. (unidentified yellow sp.1.) |
Viet Nam |
Population size unknown |
|
C. sp (unidentified yellow sp. 2.) |
Viet Nam |
Population size unknown |
|
C. edithae |
P.R. China |
Population size unknown |
|
C. grijsii |
P.R. China |
Cultivated specimen |
|
C. obtusifolia |
P.R. China |
Population size not known |
|
C. yunnanensis |
P.R. China |
Cultivated specimen |
|
C. crassissima |
P.R. China |
Population size not known |
|
C. reticulata |
P.R. China |
Population size not known |
|
C. rubrimuricata |
P.R. China |
Taxonomic status uncertain |
|
C. henryana |
P.R. China |
Cultivated specimen |
|
C. cordifolia |
P.R. China |
Cultivated specimen |
|
C. odorata |
P.R. China |
Cultivated specimen |
|
C. rhytidocarpa |
P.R. China |
Cultivated specimen |
|
C. jimpinensis |
P.R. China |
Taxonomic status uncertain |
|
C. japonica |
Viet Nam |
Cultivated specimen |
|
C. mairei |
P.R. China |
Population size not known |
* Bold print denotes newly discovered Camellia species
** Species in the process of publication, accession Nos. only
Selected morphological data derived from observations of the nine newly discovered Camellia species from Viet Nam are presented in Table 2.
Table 2. Selected morphological data for the nine, newly discovered, yellow flowering Camellia species from Viet Nam
|
Taxon |
Flower colour |
Morphology, notes |
|
C. dongnaiensis |
yellow-apricot, pink margins |
large, up to 60.0 cm long leaves |
|
C. luteocerata |
intensely yellow |
flowers laterally oblongoid |
|
C. inusitata |
light yellow |
lacks secondary branching |
|
C. sp. 0720 |
light to mid yellow, pink-lilac margins |
oblate fruit, 5.5 cm diam.
|
|
C. sp. 698 |
dark yellow |
petals in spiral arrangement |
|
C. sp. CT5 |
pale yellow |
inflorescence of 12-14 flower buds |
|
C. sp. OCa |
dark yellow |
crinkly, zigzag -like styles |
|
C. sp. OCb |
light yellow with pink blush |
filaments thickened at their base |
|
C. sp. OCc |
light yellow |
Outer filaments thin, inner wide, strap like |
Species in the process of publication, accession Nos. only
Results
Figure 1. Evolutionary phylogenies derived from the analyses of 30 multiple state morphological characters of 18 yellow flowering Camellia species using Phylip 3.69 computational program
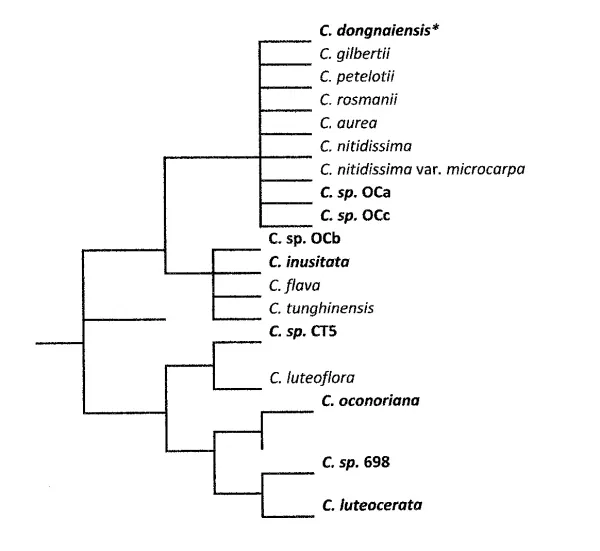
*Bold print indicates the newly discovered species
Figure 2. Interspecific relationships derived from analyses of 30 multiple state morphological characters for the nine newly discovered yellow flowering Camellia species using Phylip 3.69 computational program
 Figure 3. ISSR Consensus of 17 equal distances network for 18 Camellia species
Figure 3. ISSR Consensus of 17 equal distances network for 18 Camellia species
 Figure 4. ISSR Bootstrap
Figure 4. ISSR Bootstrap Figure 5.
Figure 5.

Discussion
The present taxonomic arrangement of genus Camellia is provisional and unsatisfactory (Chapman & Wang 2002). The existence of three valid but rather dissimilar morphology-based taxonomic systems, i.e. those of Sealy (1958), Chang & Bartholomew (1984) and Ming & Bartholomew (2007) and of at least two extensive synopses, Ho (1991) and Gao et al (2005), resulted in the taxonomy of the genus being almost unworkable. The input of molecular data, not always being in agreement with accepted morphological data, renders this situation even more challenging. It appears that the creation of a satisfactory and clearly defined classificatory system for genus Camellia is simply unattainable.
To indicate the nature of some of the issues the multiplicity of taxonomic systems presents, the authors of this paper would like to make the following comments. The majority of our studies (2003-2013) affirm the long known fact, namely the morphological closeness of already described Camellia species, yellow or otherwise (Orel et al, 2003; Orel & Marchant 2006). Figs. 1. and 2. demonstrate and interpret the existence of a number of unique morphological character dissimilarities possessed by some of the recently discovered, south Vietnamese, yellow flowering, Camellia taxa (Orel 2006; Orel & Wilson 2010b; Orel & Wilson 2012). The new yellow flowering species are morphologically so different from each other and from the known yellow flowering Camellia species (Orel et al, 2012a) that they render the existence of sect. Chrysantha Chang untenable. A substantial number of morphological and molecular characters possessed by the newly discovered yellow Camellia species do not conform to the specific parameters on which sect. Chrysantha (sensu Chang & Bartholomew) is based (Chang & Bartholomew 1984). The establishment of sect. Chrysantha Chang on the basis of petal colour and the notion of ‘systematic development routes of organ morphology’ (Chang & Ye 1994) cannot be sustained.
The above-mentioned findings (Fig. 1.) and the data presented in Fig. 2. profoundly influences the taxonomy of the yellow flowering Camellia species. The new data also influences the taxonomy of genus Camellia in general terms. The inclusion in the system of the newly discovered yellow and non-yellow flowering Camellia species (Orel & Wilson, 2010b; Orel & Wilson, 2012, Orel & Wilson, 2012b; Orel et al 2012) necessitates the widening of the terms of taxonomic reference, re genus Camellia. To achieve this, the establishment of new Camellia sections, which enabled the accommodation of the newly discovered species and their unique morphological characters, was undertaken (Orel 2006; Orel & Wilson 2010b; Orel & Wilson 2012, Orel et al, 2012a).
It should be noted that available scientific data shows that some Camellia taxa found on the Da Lat Plateau and the adjoining Lang Biang Massiff, had been subject to particular local environmental conditions, and thus are morphologically dissimilar to Camellia species of North Viet Nam and China. A divergent evolutionary process, aided by the processes of natural selection, is thus implied (Orel, G. & Marchant, A. D. 2006).
Recently obtained morphological and molecular data recommends the necessity for the retention of a number of long-established Camellia species. Good examples of this are C. nitidissima, C. tunghinensis (Figure 1.) and C. yunnanensis and C. henryana (Figures. 4. & 5.). Further, the recent morphological and molecular data (Figures 1. and 3.), show clear differences between C. petelotii, C. nitidissima and C. nitidissima var. microcarpa.
Problems with reliable identification of Camellia species are well demonstrated in Figure 3. Multiple accessions of C. petelotii and C. pingguoensis obtained from different provenances show considerable molecular dissimilarities. The authors of this work are aware of the fact that no two individual conspecific accessions are exactly the same. Calculations of genetic distances (not presented in this work), in this case, indicate dissimilarities akin to different Camellia species.
Further difficulties present themselves as the descriptions of Camellia species and the sections these are placed in vary from author to author. A good example of this is the taxonomic treatment of sect. Heterogena Sealy.
Morphological descriptors for section Heterogena, as established by Sealy (1958) were modified by Ming & Bartholomew (2007). Although similar, they are not identical. For example: Sealy (1958) - flowers…at the end of the branches, Ming & Bartholomew (2007) – flowers axillary or sub-terminal; Sealy (1958) – flowers solitary or geminate, Ming & Bartholomew (2007) – flowers solitary; Sealy (1958) – perules forming involucre, Ming & Bartholomew (2007) – …bracteoles and sepals…, no involucre is mentioned.
It should also be pointed out that the number of species and the identity of species placed within individual sections also vary from author to author. Sealy’s sect. Heterogena (1958) contains some 10 Camellia species, with C. henryana Coh. Stuart being the type species. Sect. Heterogena as re-interpreted and modified by Ming & Bartholomew (2007) contains 14 Camellia species, with the type species, C. henryana, being subsumed into C.yunnanensis var. yunnanensis.
A number of other examples may be cited, however the constraints limiting the size of this work prevent us from doing so.
Literature cited
Chang, H.T. & Bartolomew, B. 1984. Camellias. Timber Press, Portland, Oregon.
Chang, H.T. & Ye, C. 1994. Diagnosis of the systematic development of Theaceae. International Camellia Journal No. 26.
Chapman, G.P. & Wang Y.Z. 2002. The Plant Life of China. Berlin Springer.
Dellaporta, S.F., Wood, T. & Hicks, J.B. (1983) A plant DNA minipreparation: Version 2 in: Plant Molecular Biology Report 1: 19-21.
Gao, J., Parks, C.R. & Du, Y. 2005. Collected species of the genus Camellia: An illustrated outline. Zhejiang Science and Technology Press, P.R. China.
Gilmore, S., Weston, P.H. & Thomson, J.A. 1993. A simple, rapid, inexpensive and widely applicable technique for purifying plant DNA in: Systematic Botany:6: 139-148.
Ho, P-H. 1991. Cayco Viet Nam. Vol. 1., Part 1., Santa Ana, California, Mekong Printing.
Ming, T. L. & Bartholomew B. 2007. Theaceae. Pp. 366-478 in Z. Y. Wu & P. H.
Raven (editors), Flora of China, Vol. 12. Science Press, Beijing, and Missouri
Botanical Garden Press, St. Louis.
Orel, G. 2006. Notes on Camellia dongnaiensis – a new specie from Vietnam. International Camellia Journal 2006, No. 38: 90-93.
Orel, G., Marchant, A. D. & Richards, G. D. 2003. Molecular genetic investigation into evolutionary relationships of yellow-flowered Camellia species from South-East Asia. Collected Papers, International Camellia Congress 2003, March 6-March 9, Jinhua City, P. R. China.
Orel, G. & Marchant, A. D. 2006. Investigation into the evolutionary origins of Theaceae and genus Camellia. (Adapted from Melbourne Congress lecture) International Camellia Journal 2006, No. 38: 78-90.
Orel, G. & Wilson, P. G. 2010b. A New Species of Camellia Sect. Stereocarpus (Theaceae) from Vietnam. - Novon 20: 198-202.
Orel, G. & Wilson, P. G. 2012. Camellia cattienensis: a new species of Camellia (sect. Archaecamellia: Theaceae) from Viet Nam. Kew Bulletin 66: 565-569.
Orel, G., Wilson, P. G., Curry, A. S. & Luu Hong Truong (2012a). Camellia inusitata (Theaceae), a new species forming a new section (Bidoupia) from Vietnam.
Edinburgh Journal of Botany 69: 347-355.
Orel, G., Wilson, P. (2012b). Camellia cherryana (Theaceae), a new species from China. Annales Botanici Fennici 49: 248-254.
Orel, G., Wilson, P. G., Curry, A. S. & Luu Hong Truong (2013). Camellia oconoriana (Theaceae) a new species from Vietnam. Edinburgh Journal of Botany 70: 439-447
Sealy, J. R. 1958. A revision of the genus Camellia. Royal Horticultural Society.
Singh M, Bandana and Ahuja PS, 1999. Isolation and PCR amplification of genomic DNA from market samples of dry tea. Plant Molecular Biology Reporter 17: 171-178.
Wilkie, S. 1997. Isolation of total genomic DNA. Plant molecular biology-a
Laboratory Manual. London: Springer.
Web design by Tribal Systems